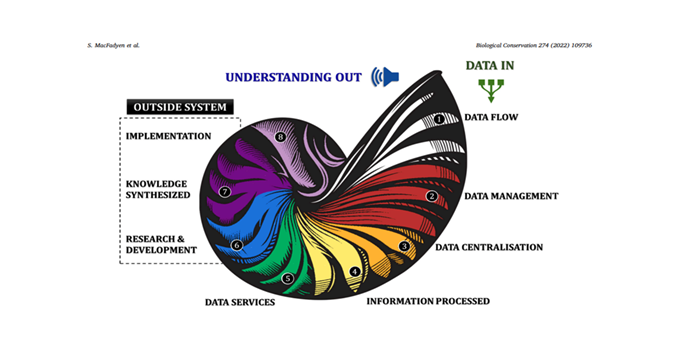



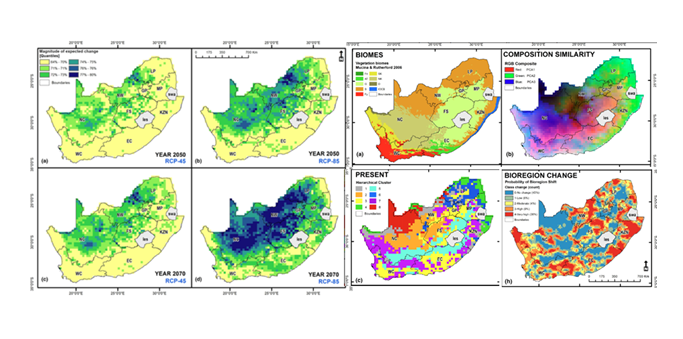

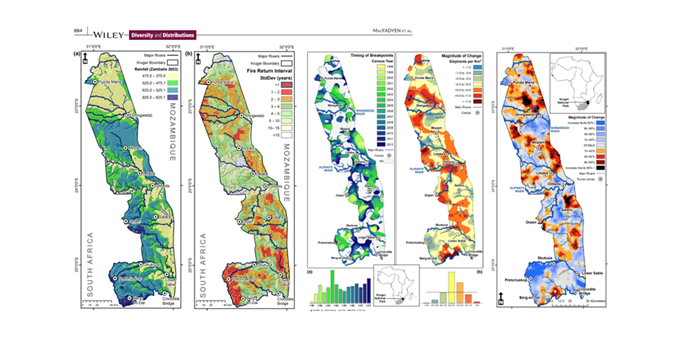

Cross-Scale Consistency in Biodiversity Patterns
Ecological patterns are emerging structures observed in populations, communities and ecosystems. Elucidating drivers behind ecological patterns can greatly improve our knowledge on how ecosystems assemble, function and respond to change and perturbation. Due to the non-random nature, most, if not all, ecological patterns change with measurement and organization scales and exhibit distinct scaling properties. Models in this section are dealing with inferring patterns across scales that are in close proximity. As such, these models are often developed to ensure the consistency of measurements across different scales. To ensure the consistency, patterns across scales are normally bridged using probability theory. Specifically, this section presents models that investigate how aggregated structures of organisms and biodiversity change with measurement scales, which biological patterns resonate with underlying processes at the same characteristic scales, and why.
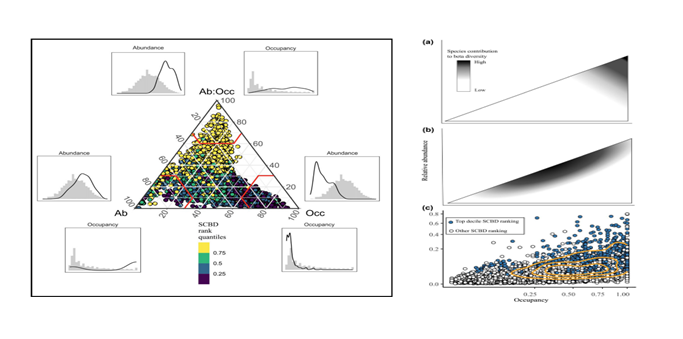
Scaling pattern of occupancy
Species distributions are not uniform across space, reflecting the interplay between habitat heterogeneity and the underlying nonlinear biotic regulation. When ecologists examine such non-random, aggregated patterns across scales, the Modifiable Areal Unit Problem presents itself. The problem can be described as the change in species distribution characteristics as the grain and extent of sampling change. The aim here is to depict how species occupancy and its aggregation level change when scaling up and down, and therefore provide a universal basis for ensuring cross-scale consistency. Under certain conditions, these models further allow extrapolating fine-scale occupancy and population densities from coarse-scale observations. Great potential exists to apply such predictive models in various cross-scale pattern analyses.
Species turnover and co-occurrence
Species diversity patterns, such as the species-area curve, endemics-area relationship, distance decay of similarity and occupancy frequency distribution, reflect the scale-dependence of species turnover. Measures of spatial turnover in the compositional similarity or difference between assemblages are commonly based on beta diversity which was originally derived for pairwise comparisons of individual assemblages. However, none of the metrics of species turnover are able to fully reduce all diversity partitions in multiple-assemblage cases, i.e. the diversity components of three or more assemblages cannot be completely expressed using pairwise species turnover. That is, pairwise metrics are not entirely adequate for depicting compositional similarity across multiple assemblages. To exploit resources while mitigating conflicts, species often partition available habitats, forming co-distribution patterns of association or dissociation. Null models based on permutation or neutral/niche processes have been widely applied for detecting signals of bio-interactions from co-occurrence patterns. However, co-occurrence is scale dependent and should be used with caution for inferring biotic interactions or processes.
Space-for-time substitution
The spatial and temporal scales of ecological processes are intertwined. Processes that account for the spatial distribution of species also underpin its temporal dynamics. This means that we can potentially forecast the future or rebuild the history based solely on current spatial distribution, without resorting to long-term time series. Specifically, the scaling pattern of occupancy has been found to be related to population trends. As the ability to forecast the temporal trend of a focal species provides crucial information on its performance and viability, the methodology of space-for-time substitution is extremely appealing, especially since our ability to obtain spatial records has been drastically improved in recent years.
Scale resonance
Just as two tuning forks of the same characteristic frequency resonate, so do ecological patterns and processes working at the same scale. Species distributions are regulated by a variety of abiotic and biotic processes working in concert but at different characteristic scales, whereas identified key processes, e.g. using multivariate statistics, are those resonate with the measurement scale of the study. That is, information being picked up is diluted by the measurement scale, rather than the intrinsic cross-scale mechanism. This finding brings into question many regional management planning practices that are based on the upscale extrapolation of local-scale studies.
Community assembly rules
The two dominant theories on the development and structure of communities are niche and neutral theory. Niche theory explains the structure of communities using the relationship between species traits and habitat characteristics. Meanwhile, neutral theory assumes a fixed species pool in the absence of speciation and invasion, and considers all species to be ecologically equivalent, with stochastic dispersal and ecological drift being the only processes determining community structure. Despite contrasting opinions on the value of neutral theory, it is now generally accepted that neutral and niche processes interact in natural communities and both contribute towards the structure of species assemblages. The relative roles of neutral and niche processes have been shown to differ across spatio-temporal scales and modelling these processes in combination better represents biological patterns than neutral or niche models alone.